
Sendero: An Extended, Agent-Based Implementation of Kauffman's NKCS Model
Journal of Artificial Societies and Social Simulation
12 (4) 8
<https://www.jasss.org/12/4/8.html>
For information about citing this article, click here
Received: 12-Oct-2008 Accepted: 21-Jun-2009 Published: 31-Oct-2009

 Abstract
Abstract

|
| Figure 1. Mathematical formalization of the NK model |
We will treat each species as though its entire population were genetically identical. At each "generation", the population will seek a fitter genotype by mutating a single, randomly chosen gene to the alternative allele. If the new mutant genotype is fitter, the population will move to this new point on its landscape. (Kauffman 1995, pp. 225-226)
|
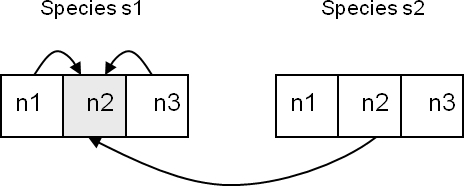
| Figure 2. Gene n2 in species s1 depends on n1 and n3 (K = 2) and n2 from species s2 (C = 1) |
|
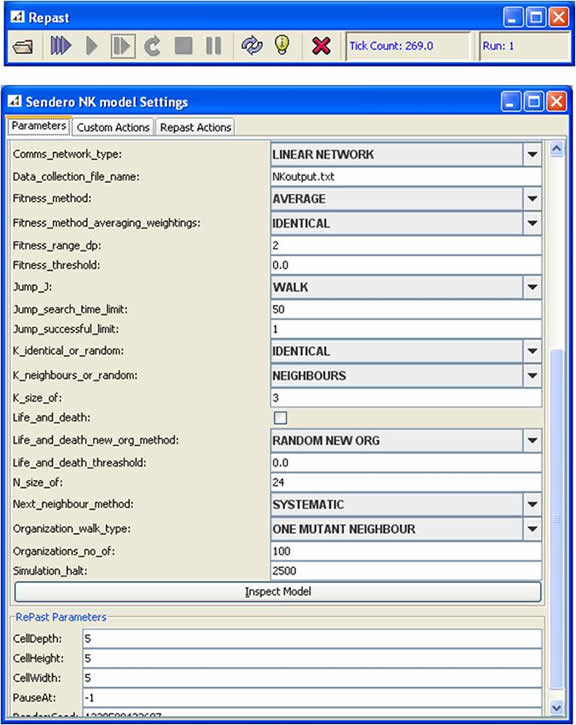
| Figure 3. The Sendero parameter interface (generated by Repast) |
|
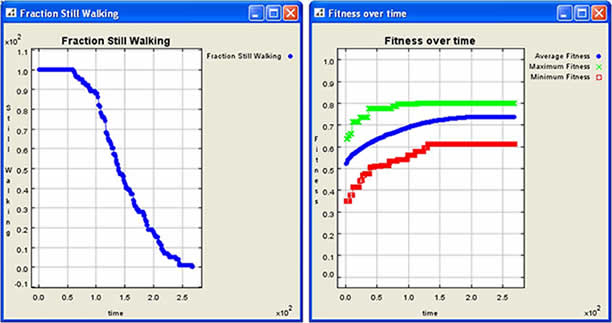
| Figure 4. Sample graphical output for Sendero (generated by Repast) |
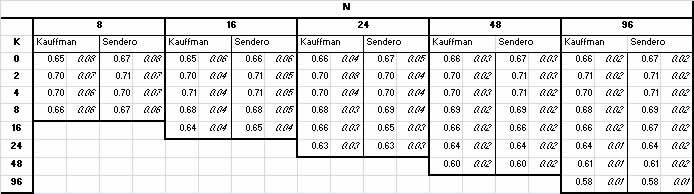 |
| Table 1. Mean fitness of local optima - Kauffman's Table 2.1, p. 55 results compared with Sendero. Note: along the diagonal where K = N, actual value is K - 1 |
|
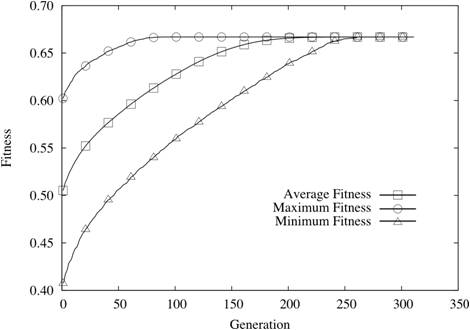
| Figure 5. N = 24 and K = 0 |
|
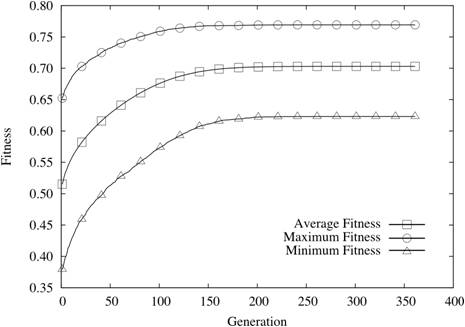
| Figure 6. N = 24 and K = 4 |
|
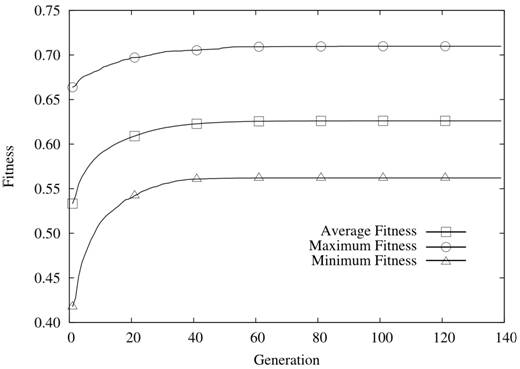
| Figure 7. N = 24 and K = 23 |
|
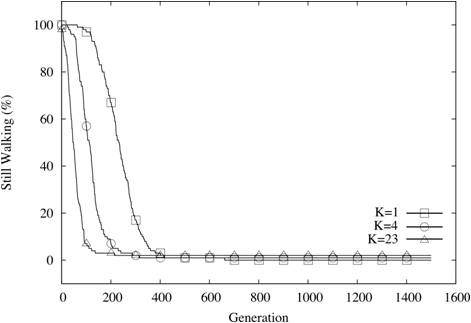
Video 1. Shows a population of 100 agents, each with 48 alleles having
epistatic interactions with all 47 others, leading to a maximally
rugged fitness landscape. Average fitness quickly reaches a local
maximum. (Click on the movie for a larger version in a new window) |
|
| Figure 8. Coevolutionary sets still walking (N=24, C=1, K varies) |
|
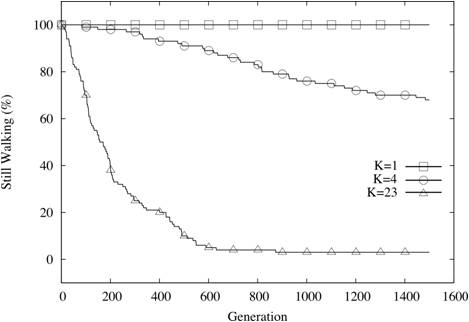
| Figure 9. Coevolutionary sets still walking (N=24, C=8, K varies) |
|
Video 2. Shows a population of five co-evolutionary sets, each comprising
five species. The fitness of each species within the co-evolutionary
set is dependent on interactions with two other species in the set.
Fitness of the sets proceeds chaotically at first, before resolving to
a stable maximum after 450 generations. (Click on the movie for a larger version in a new window) |
|
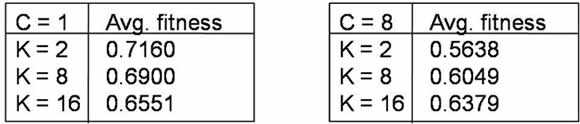
| Table 2. Final fitness values for NKCS simulations shown in Figures 7 and 8 |
EHRLICH, P. R. and P. H.. Raven, (1964). Butterflies and Plants: A Study in Coevolution. Evolution, 18(4): 586-608.
JERMAIS, J., and L. Gani, (2004). Integrating business strategy, organizational configurations and management accounting systems with business unit effectiveness: a fitness landscape approach. Management Accounting Research, 15: 179-200.
KAUFFMAN, S., (1993). The Origins of Order: Self-Organization and Selection in Evolution. Oxford University Press.
KAUFFMAN, S., (1995). At Home In The Universe. Oxford University Press.
LAZER, D., and A. Friedman, (2007). The social structure of exploration and exploitation. Administrative Science Quarterly 52: 667-694.
LEVINTHAL, D. A., (1997). Adaptation on rugged landscapes. Management Science, 43(7): 934-950.
LEVITAN, B., J. Lobo, R. Schuler, and S. Kauffman, (1997). Evolution of organizational performance and stability in a stochastic environment. Computational & Mathematical Organization Theory, 8: 281-313.
MCCARTHY, I. P., (2002). Manufacturing fitness and NK models. In G. Frizelle and H Richards, editors, Tackling Industrial Complexity. Institute for Manufacturing, Cambridge, UK. Available via www.ifm.eng.cam.ac.uk/mcn/proceedings.htm. Retrieved January 2008.
MERZ, P., (2000). Memetic Algorithms for Combinatorial Optimization Problems: Fitness Landscapes and Effective Search Strategies. PhD thesis, University of Siegen.
NORTH, M., N. T. Collier, and J. R. Vos, (2006). Experiences creating three implementations of the repast agent modeling toolkit. ACM Trans. Model. Comput. Simul., 16(1): 1-25.
PADGET, J., and R. T. Vidgen, (2008). The Sendero Project. On-line access at http://wiki.bath.ac.uk/display/sendero. Retrieved October 2008.
RIVKIN, J., (2000). Imitation of complex strategies. Management Science, 46(6): 824-844.
WEINBERGER, E., (1996). NP Completeness of Kauffman's N-k Model, a Tuneably Rugged Fitness Landscape. Sante Fe Institute working paper 96-02-003. Available via http://www.santafe.edu/research/publications/workingpapers/96-02-003.ps. Retrieved 20090313.
WRIGHT, S., (1932). The role of mutation, inbreeding, crossbreeding and selection in evolution. In Proceedings of the Sixth International Congress on Genetics, volume 1, pages 356-366.
YUAN, Y., and B. McKelvey, (2004). Situated Learning Theory: Adding Rate and Complexity Effects via Kauffman's NK Model. Nonlinear Dynamics, Psychology, and Life Sciences, 8: 65-102.
 Return to Contents of this issue
Return to Contents of this issue
© Copyright Journal of Artificial Societies and Social Simulation, [2009]